From Proteopedia
proteopedia linkproteopedia link
Introduction
MR1 is an antigen presenting molecule, related to major histocompatibility complex class I (MHC-I). MR1 primary structure is highly conserved across mammalian species, suggesting an evolutionarily conserved function for MR1 in immunity.[1] This molecule, when bound and presenting its ligand, specifically activates mucosal-associated invariant T (MAIT) cells, a subset of the innate-like T-cell population.[2] It has been found that MR1 ideally binds ligands originating from vitamin metabolites, particularly those derived from microbial riboflavin (vitamin B2) biosynthetic pathways.[3] Since there are several vitamin biosynthetic pathways that are unique to some species of bacteria or yeast, it is possible that MAIT cells use these metabolites (via MR1) to detect microbial infection.
MR1 is MHC-I Related
MHC-I consists of two chains: a large alpha chain and a smaller beta chain. The beta chain is called β2-microglobulin (β2m), which stabilizes the entire complex when MHC-I binds its ligand. β2m is linked to the alpha chain noncovalently via the α3 domain of the alpha chain. MHC-I binds peptides generated by degradation of cytosolic proteins by cellular proteosomes. These peptides must be transported from the cytosol into the endoplasmic reticulum (ER) in order to meet the MHC-I molecule, whose peptide binding site is faced towards the lumen of the ER. The MHC-I-peptide complex then leaves the ER through the secretory pathway to become inserted in the plasma membrane of the cell. If the presented peptide is foreign, and therefore possibly pathogenic, cytotoxic T cells will recognize the complex and kill the presenting cell.
The α3 domain of MHC-I is the transmembrane component of the complex. It interacts with β2m and with CD8 co-receptors of T cells. The main binding site for CD8 are residues 223-229 of the alpha chain. The α1 and α2 domains form a binding groove for peptides. This groove has both deep and shallow pockets that accommodate the side chains of specific peptides.
Similarly the MHC-I, MR1 also consists of an alpha chain and β2m. MR1's α1 and α2 domains also serve as the molecule's antigen-binding cleft. However, the structure and chemistry of this site does not accomodate peptide or lipid-based antigens.
Structure of MR1-antigen Complex
| Recovery of properly assembled MR1-β2m complexes indicates efficient capture of a ligand. Using this approach, it was found that MR1 successfully refolded in the presence of folic acid. Further analysis of the complex revealed that 6-formyl pterin (6-FP) was the specific ligand component that allowed proper refolding of MR1.
|
MR1-restricted MAIT Activation
MAIT cells compose up to 10% of the peripheral blood T-cell population in humans. They are mostly associated with mesenteric lymph nodes and the gastrointestinal mucosa.
[4] They interact with the MR1-antigen complex via their αβ T-cell antigen receptor (TCR). These TCRs consist of an α and β chain, and each chain has a constant (C) and variable (V) domains.
[5] The V domain of each chain has three complimentarity determining regions (CDRs). Together, the six CDRs form a ligand-binding site for the MAIT TCR. The MAIT TCR is restricted to MR1-antigen presentation. Mutational data suggests that a limited number of Vα chain residues of the MAIT TCR are critical for MR1-induced activation.
[6]
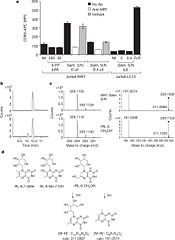
Identification of bacterially-derived MAIT-cell antigens
Even though 6-FP is a ligand for MR1, it does not activate MAIT cells. It was found that antigen-presenting cells transfected with MR1 and infected with Salmonella typhimurium specifically activated MAIT cells. Notably, the supernatent from S. typhimurium was able to activate MAIT cells. Activation was assayed by CD69 upregulation and intracellular cytokine staining for interferon (IFN)-γ and tumor necrosis factor (TFN).[7] By searching for ligands from the supernatent that complexed with MR1, it was revealed that candidate MAIT-activating ligands are derivatives of riboflavin (vitamin B2).[8] These vitamin metabolites are structurally close to 6-FP, but they possess an extra ribityl moiety that may permit direct contact by the MAIT TCR. These compounds are derived from the riboflavin biosynthetic pathway present in most, but not all, bacteria and yeast.
Biological Significance
There are a wide range of microbes that stimulate MAIT cells via MR1 interaction, and they all synthesize riboflavin. This suggests a possible mode by which MAIT cells might sense microbial infection or overgrowth at mucosal sites in an MR1-restricted manner.[9] There are possibly many species of bacteria in the gut flora that have riboflavin biosynthetic pathways. Perhaps the relatively frequent stimulation of MAIT cells serves as way for the body to strengthen the immune system, so as to not be overcome by the co-existing microbiota. The human microbiome is very delicately balanced. It is possible that a spiked growth of an opportunistic pathogen could trigger extensive MAIT cell activation (via MR1) to cause an effective immune response against the pathogen. There also could be other microbial-specific metabolites[10]that may serve as indicators of microbial infections that undergo immunosurveillance.
The pterin ring, an important component of MR1 ligands, occurs widely in nature and also represents a common scaffold of small molecule therapeutics.[11] Further research must be done to determine if MAIT cell-MR1 interactions are affected by drugs and diet.
References
- ↑ Huang S, Martin E, Kim S, Yu L, Soudais C, Fremont DH, Lantz O, Hansen TH. MR1 antigen presentation to mucosal-associated invariant T cells was highly conserved in evolution. Proc Natl Acad Sci U S A. 2009 May 19;106(20):8290-5. Epub 2009 Apr 30. PMID:19416870 doi:10.1073/pnas.0903196106
- ↑ Huang S, Gilfillan S, Cella M, Miley MJ, Lantz O, Lybarger L, Fremont DH, Hansen TH. Evidence for MR1 antigen presentation to mucosal-associated invariant T cells. J Biol Chem. 2005 Jun 3;280(22):21183-93. Epub 2005 Mar 31. PMID:15802267 doi:10.1074/jbc.M501087200
- ↑ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature. 2012 Oct 10. doi: 10.1038/nature11605. PMID:23051753 doi:10.1038/nature11605
- ↑ Treiner E, Duban L, Bahram S, Radosavljevic M, Wanner V, Tilloy F, Affaticati P, Gilfillan S, Lantz O. Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature. 2003 Mar 13;422(6928):164-9. PMID:12634786 doi:10.1038/nature01433
- ↑ Reantragoon R, Kjer-Nielsen L, Patel O, Chen Z, Illing PT, Bhati M, Kostenko L, Bharadwaj M, Meehan B, Hansen TH, Godfrey DI, Rossjohn J, McCluskey J. Structural insight into MR1-mediated recognition of the mucosal associated invariant T cell receptor. J Exp Med. 2012 Mar 12. PMID:22412157 doi:10.1084/jem.20112095
- ↑ Reantragoon R, Kjer-Nielsen L, Patel O, Chen Z, Illing PT, Bhati M, Kostenko L, Bharadwaj M, Meehan B, Hansen TH, Godfrey DI, Rossjohn J, McCluskey J. Structural insight into MR1-mediated recognition of the mucosal associated invariant T cell receptor. J Exp Med. 2012 Mar 12. PMID:22412157 doi:10.1084/jem.20112095
- ↑ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature. 2012 Oct 10. doi: 10.1038/nature11605. PMID:23051753 doi:10.1038/nature11605
- ↑ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature. 2012 Oct 10. doi: 10.1038/nature11605. PMID:23051753 doi:10.1038/nature11605
- ↑ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature. 2012 Oct 10. doi: 10.1038/nature11605. PMID:23051753 doi:10.1038/nature11605
- ↑ Nicholson JK, Holmes E, Kinross J, Burcelin R, Gibson G, Jia W, Pettersson S. Host-gut microbiota metabolic interactions. Science. 2012 Jun 8;336(6086):1262-7. Epub 2012 Jun 6. PMID:22674330 doi:10.1126/science.1223813
- ↑ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature. 2012 Oct 10. doi: 10.1038/nature11605. PMID:23051753 doi:10.1038/nature11605

