User:Joseph Lipsick/RAS
From Proteopedia
|
|
|
The RAS gene family was discovered due to the presence of two closely related retroviral cancer genes (oncogenes)
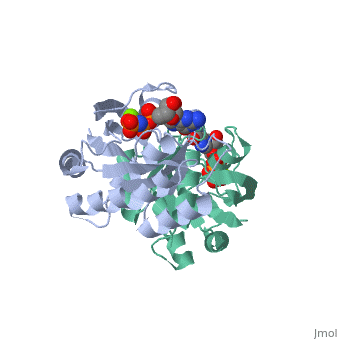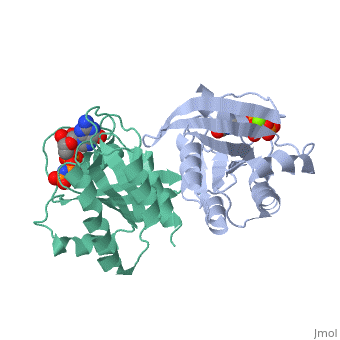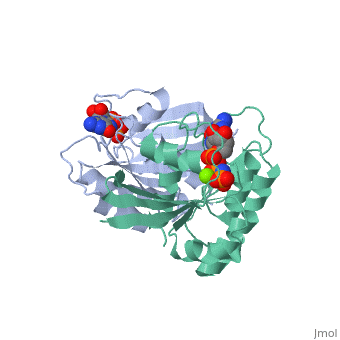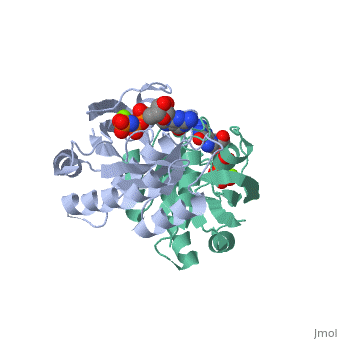
within the Harvey and Kirsten RAt Sarcoma viruses. These retroviral oncogenes arose via capture of two normal cellular genes, H-RAS and K-RAS. A DNA transfection assay for oncogenes in human bladder cancer later identified the same H-RAS gene. Remarkably, the mutated oncogenic form of H-RAS differed from its normal counterpart by a single nucleotide change that caused a single amino acid substitution. Biochemical studies of purified RAS proteins revealed that they were capable of binding nucleotides, with a particularly high affinity for GTP. Extensive genetic, biochemical, and structural studies have established a model in which the RAS proteins function as molecular switches, as do related GTP-binding proteins. The RAS switch is "ON" when it binds , and is "OFF" when it binds . The changes in protein conformation between the and states are not very large. These changes primarily occur in two regions known as SWITCH I (yellow) and SWITCH II (magenta), and can be visualized by toggling the spin off and on in these partial space-filling models. The RAS protein itself has an intrinsic GTPase activity, thereby limiting the duration of time spent in the "ON" configuration. The two most common oncogenic mutations in H-RAS affect residues and , both of which are adjacent to the bound GTP molecule. These oncogenic mutations greatly inhibit the intrinsic GTPase activity, thereby causing the RAS switch to spend more time in the "ON" position. The RAS proteins are present at the plasma membrane and transmit signals from transmembrance receptor tyrosine kinases (e.g. EGF and PDGF receptors) to downstream intracellular effectors that include the MAPK protein kinase cascade and the PI3K lipid kinase. Binding of EGF or PDGF to their receptors causes a relocalization of a Guanine Nucleotide Exchange Factor (GEF) protein to the plasma membrane. The structure of , a prototypic GEF, together with RAS implies that the GEF prys open the nucleotide binding site with a loss of bound GDP. The ten-fold higher ratio of GTP to GDP within the cell results in the replacement of RAS-GDP ("OFF" state) with RAS-GTP ("ON" state). Consistent with this model, gain-of-function mutations of GEF genes can be oncogenic (e.g. VAV oncogene) even in the absence of mutations of RAS. Conversely, a GTPase Activating Protein () can bind to RAS-GTP and increase the rate of the intrinsic RAS GTPase. Consistent with this model, loss-of-function mutations of GAP genes (e.g. NF1 tumor suppressor) can be oncogenic even in the absence of mutations of RAS itself.
Milburn, et al. Science. 1990. 247: 939-45.
Wittinghofer and Pai. Trends Biochem Sci. 1991. 16: 382-7.
Malumbres and Barbacid. Nature Rev Cancer. 2003. 3: 459-65.
Bos, Rehmann, and Wittinghofer. Cell. 2007. 129: 865-77.
Karnoub and Weinberg. Nature Rev Mol Cell Biol. 2008. 9: 517-531.
