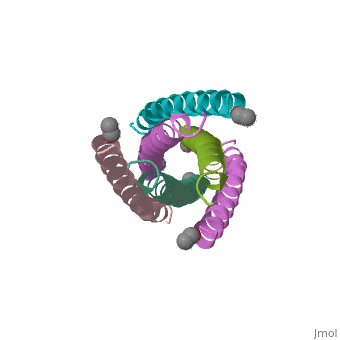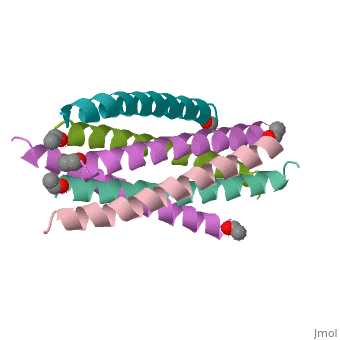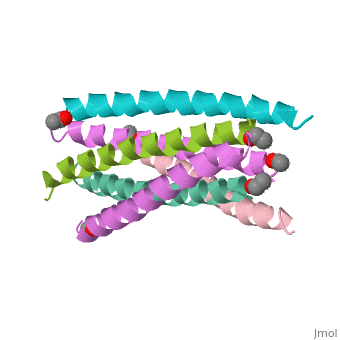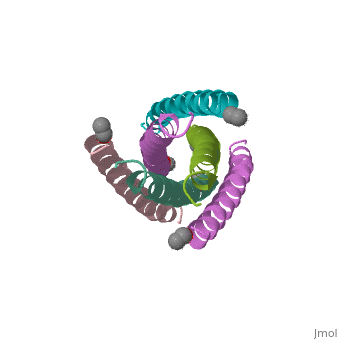1aik
From Proteopedia
HIV GP41 CORE STRUCTURE
Structural highlights
FunctionENV_HV1H2 The surface protein gp120 (SU) attaches the virus to the host lymphoid cell by binding to the primary receptor CD4. This interaction induces a structural rearrangement creating a high affinity binding site for a chemokine coreceptor like CXCR4 and/or CCR5. This peculiar 2 stage receptor-interaction strategy allows gp120 to maintain the highly conserved coreceptor-binding site in a cryptic conformation, protected from neutralizing antibodies. Since CD4 also displays a binding site for the disulfide-isomerase P4HB/PDI, a P4HB/PDI-CD4-CXCR4-gp120 complex may form. In that complex, P4HB/PDI could reach and reduce gp120 disulfide bonds, causing major conformational changes in gp120. TXN, another PDI family member could also be involved in disulfide rearrangements in Env during fusion. These changes are transmitted to the transmembrane protein gp41 and are thought to activate its fusogenic potential by unmasking its fusion peptide.[1] Surface protein gp120 (SU) may target the virus to gut-associated lymphoid tissue (GALT) by binding host ITGA4/ITGB7 (alpha-4/beta-7 integrins), a complex that mediates T-cell migration to the GALT. Interaction between gp120 and ITGA4/ITGB7 would allow the virus to enter GALT early in the infection, infecting and killing most of GALT's resting CD4+ T-cells. This T-cell depletion is believed to be the major insult to the host immune system leading to AIDS (By similarity).[2] The surface protein gp120 is a ligand for CD209/DC-SIGN and CLEC4M/DC-SIGNR, which are respectively found on dendritic cells (DCs), and on endothelial cells of liver sinusoids and lymph node sinuses. These interactions allow capture of viral particles at mucosal surfaces by these cells and subsequent transmission to permissive cells. DCs are professional antigen presenting cells, critical for host immunity by inducing specific immune responses against a broad variety of pathogens. They act as sentinels in various tissues where they take up antigen, process it, and present it to T-cells following migration to lymphoid organs. HIV subverts the migration properties of dendritic cells to gain access to CD4+ T-cells in lymph nodes. Virus transmission to permissive T-cells occurs either in trans (without DCs infection, through viral capture and transmission), or in cis (following DCs productive infection, through the usual CD4-gp120 interaction), thereby inducing a robust infection. In trans infection, bound virions remain infectious over days and it is proposed that they are not degraded, but protected in non-lysosomal acidic organelles within the DCs close to the cell membrane thus contributing to the viral infectious potential during DCs' migration from the periphery to the lymphoid tissues. On arrival at lymphoid tissues, intact virions recycle back to DCs' cell surface allowing virus transmission to CD4+ T-cells. Virion capture also seems to lead to MHC-II-restricted viral antigen presentation, and probably to the activation of HIV-specific CD4+ cells.[3] The transmembrane protein gp41 (TM) acts as a class I viral fusion protein. Under the current model, the protein has at least 3 conformational states: pre-fusion native state, pre-hairpin intermediate state, and post-fusion hairpin state. During fusion of viral and target intracellular membranes, the coiled coil regions (heptad repeats) assume a trimer-of-hairpins structure, positioning the fusion peptide in close proximity to the C-terminal region of the ectodomain. The formation of this structure appears to drive apposition and subsequent fusion of viral and target cell membranes. Complete fusion occurs in host cell endosomes and is dynamin-dependent, however some lipid transfer might occur at the plasma membrane. The virus undergoes clathrin-dependent internalization long before endosomal fusion, thus minimizing the surface exposure of conserved viral epitopes during fusion and reducing the efficacy of inhibitors targeting these epitopes. Membranes fusion leads to delivery of the nucleocapsid into the cytoplasm.[4] The envelope glyprotein gp160 precursor down-modulates cell surface CD4 antigen by interacting with it in the endoplasmic reticulum and blocking its transport to the cell surface (By similarity).[5] The gp120-gp41 heterodimer seems to contribute to T-cell depletion during HIV-1 infection. The envelope glycoproteins expressed on the surface of infected cells induce apoptosis through an interaction with uninfected cells expressing the receptor (CD4) and the coreceptors CXCR4 or CCR5. This type of bystander killing may be obtained by at least three distinct mechanisms. First, the interaction between the 2 cells can induce cellular fusion followed by nuclear fusion within the syncytium. Syncytia are condemned to die from apoptosis. Second, the 2 interacting cells may not fuse entirely and simply exchange plasma membrane lipids, after a sort of hemifusion process, followed by rapid death. Third, it is possible that virus-infected cells, on the point of undergoing apoptosis, fuse with CD4-expressing cells, in which case apoptosis is rapidly transmitted from one cell to the other and thus occurs in a sort of contagious fashion.[6] The gp120-gp41 heterodimer allows rapid transcytosis of the virus through CD4 negative cells such as simple epithelial monolayers of the intestinal, rectal and endocervical epithelial barriers. Both gp120 and gp41 specifically recognize glycosphingolipids galactosyl-ceramide (GalCer) or 3' sulfo-galactosyl-ceramide (GalS) present in the lipid rafts structures of epithelial cells. Binding to these alternative receptors allows the rapid transcytosis of the virus through the epithelial cells. This transcytotic vesicle-mediated transport of virions from the apical side to the basolateral side of the epithelial cells does not involve infection of the cells themselves.[7] Evolutionary Conservation Check, as determined by ConSurfDB. You may read the explanation of the method and the full data available from ConSurf. Publication Abstract from PubMedThe envelope glycoprotein of human immunodeficiency virus type 1 (HIV-1) consists of a complex of gp120 and gp41. gp120 determines viral tropism by binding to target-cell receptors, while gp41 mediates fusion between viral and cellular membranes. Previous studies identified an alpha-helical domain within gp41 composed of a trimer of two interacting peptides. The crystal structure of this complex, composed of the peptides N36 and C34, is a six-helical bundle. Three N36 helices form an interior, parallel coiled-coil trimer, while three C34 helices pack in an oblique, antiparallel manner into highly conserved, hydrophobic grooves on the surface of this trimer. This structure shows striking similarity to the low-pH-induced conformation of influenza hemagglutinin and likely represents the core of fusion-active gp41. Avenues for the design/discovery of small-molecule inhibitors of HIV infection are directly suggested by this structure. Core structure of gp41 from the HIV envelope glycoprotein.,Chan DC, Fass D, Berger JM, Kim PS Cell. 1997 Apr 18;89(2):263-73. PMID:9108481[8] From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine. See AlsoReferences
| ||||||||||||||||||||